Research

Structure and Function of Macromolecular Machines
The general goal of the Schmeing lab is to understand how large, natural biosynthetic enzymes make their interesting and important products. The natural products are typically secondary metabolites, which are not essential for life, but which give the organisms that make them an advantage in their complex environments. Two of our favourite types of biosynthetic enzymes are nonribosomal peptide synthetases (NRPS), which synthesize compounds like the antibiotic daptomycin, the anti-tumour actinomycin D and the immunosuppressant cyclosporin, and cyanophycin synthetase, which makes an extremely useful green polymer. We combine cryo-EM, crystallography, biophysical and biochemical techniques to visualize and understand these awesome enzymes.
Nonribosomal Peptide Synthetases
Nonribosomal peptide synthetases (NRPS) are large macromolecular machines that, like the ribosome, catalyze peptide bond formation. Instead of making proteins, these enzymes produce a large variety of small molecules with important and diverse biological activity.

NRPS amide bond forming domains
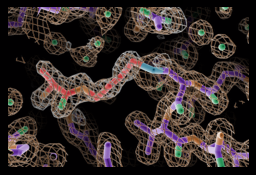
In basic NRPS modules, the C domain catalyzes the key catalytic event of NRPS function, amide bond formation to link substrate monomers and grow the peptide chain. The C domain is a pseudodimer of ~450 amino acids, with N- and C-terminal subdomains. The active site includes a HHxxxDG sequence motif that sits at the bottom of a covered “canyon” formed by the two subdomains. With our first crystal structure solved in the lab, we described important movements that occur between the subdomains that may facilitate C domain catalysis.
NRPS tailoring domains
Tailoring domains in NRPSs allow products to access far more chemical space and are commonly found in these synthetases. For example, cyclosporin synthetase contains methyltransferase domains; daptomycin (Cubicin) synthetase, epimerization domains; bactitracin (BACiiM) synthetase, a heterocyclization domain.
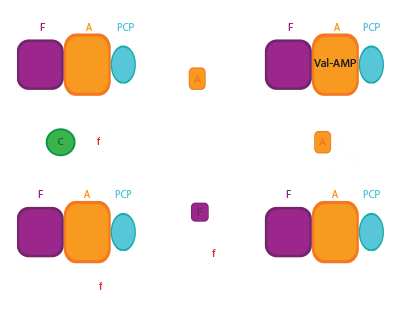

NRPS termination domains
NRPSs need specialized termination domains to release the final product. This is often the thioesterase (TE) domain. The TE catalytic mechanism proceeds through two half reactions: In the first step, the linear substrate made by the enzyme is transferred from the final PCP domain to the TE domain active site serine, forming a covalently attached acyl-enzyme intermediate.
Large NRPS proteins
In their assembly-line logic and complicated catalytic cycle, NRPS domains and modules must work in concert to synthesize the nonribosomal peptide product. To investigate the holistic workings of NRPSs, we use X-ray crystallography, chemical biology tools, biophysical techniques and biochemical methods to elucidate their structures and functions.

Cyanophycin synthesis and degradation
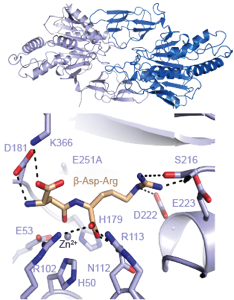
The ribosome (past research)
The ribosome is the cell’s protein factory. It translates the genetic information in mRNA into protein, rapidly and with high fidelity, using aminoacyl-tRNAs as substrates. A large number of accessory protein factors are necessary for in vivo protein synthesis, and the interplay between these factors and the ribosome is extremely complex. Deregulation of protein synthesis in humans in associated with cancers, and many important antibiotics target the bacterial ribosome.